Experientia 15(4): 190-199 (Apr 15, 1964)
Polygenic Sex Determination
By CURT KOSSWlG
pp. 192-193
In chironomids it was found that a certain segment attached to a given chromosome is the carrier of male-determining genes (BEERMANN14). This segment can be attached either to one or the other of the haploid set of chromosomes of these forms. The chromosome which bears the segment in question becomes the Y-chromosome. Only one male-determining section can be present in order to assure the development of a male. Therefore, even in the same population, one type of female X1X1X2X2AA, but two types of males, X1Y1X2X2AA or X1X1X2Y2AA, are possible. There are good reasons to believe that the M-genes in these forms are localized in the transportable section, whereas we have no knowledge about the localization of female-determining genes in this case. F-genes may be situated in the X's, in the autosomes, or in both; a decision between these possibilities is difficult. Nevertheless it seems to be true that, in the case of Chironomus, a single dose of M-genes in a transferable chromosomal segment is stronger in its phenotypical effect than the sum of F-genes in the double set of autosomes and/or X's. A somewhat similar situation was observed in some Cyprinodontid fishes, e.g. in Lebistes (WINGE15,16) and in Platypoecilus (GORDON6,17). It is highly probable that a segment of the Y, non-homologous to any section of the X, bears the realizator for the heterogametic sex18. In Lebistes this is normally the male; in Platypoecilus maculatus it is either the male or the female depending on the geographic position of the population. In the case of male heterogamety, it could be proved for Platypoecilus that the X-chromosomes bear female determiners, the M-gene (or a complex of them) being localized in a specialized segment of the Y. But in the autosomes, too, there are genes contributing to sex-determination. ÖKTAY19,20 was able to show that in an incest-line of Platypoecilus maculatus from Mexico, males of the constitution XX occurred. Until now, ÖKTAY has succeeded in producing such exceptional XX-males by brother-sister matings during 9 generations (Figure 1). The hereditary basis of this type of male production is beyond any doubt. Outcrossing of XX-exceptional males to normal XX-females results practically only in female progeny. On the other hand, interbreeding of males and females of the exceptional line led to both sexes in extremely variable proportions in different crossings.
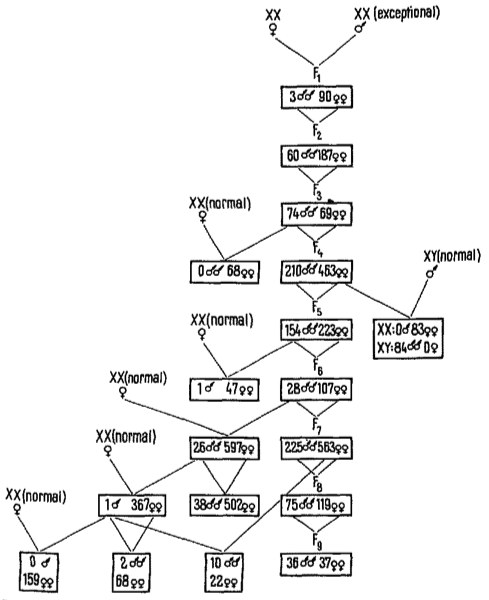 |
| Fig, 1. Schematic representation of ÖKTAY'S experiments with a Platypoecilus maculatus form showing polygenic sex determination. Outcrossing with the normal stock results practically in the restitution of the monogenic sex determination type. |
The situation in ÖKTAY'S exceptional line finds a satisfying interpretation on the following basis. The presence of at least a certain number of M-genes, localized in different autosomes, has to be assured by a recombination effect during meiosis and zygote formation. Only those individuals which bear so many M-genes that the threshold for male differentiation is reached become males. Other combinations without M-genes (if there are any) or with a number below the threshold value will differentiate into females. Naturally it is also possible, and to be expected, that such males will be produced which bear more M-genes than necessary for reaching the threshold. The latter may be called strong males. The strength of a male cannot be recognized in its phenotype. All males reach sexual maturity approximately at the same age and at the same body-length. Strong and weak males can be distinguished from one another only by the number of male offspring they produce with a given female. The strong male will have a higher percentage of male offspring than a weak one. In practice such an experiment is not easily performed because Platypoecilus is a viviparous fish, one copulation being sufficient for a number of broods. As will be mentioned later, the correctness of this view could be proved using (BREIDER21) 'marked' spermatozoa (bearing dominant colour genes) in Xiphophorus helleri, which in general has this type of polygenic sex-determination resulting from a more or less high number of freely combinable genes localized in its 'autosomes' (Table I and II). This is why, in extending the exceptional line of her XX-Platys, ÖKTAY is obliged to work somewhat 'in the dark', as neither the strength of the male used in a certain cross is known nor the number of M-genes below the threshold-line is to be recognized from the phenotype of a female. A weak female will produce a higher percentage of males than a strong one, when the two are coupled with the same male, whatever the strength of the latter may be. This experiment has been performed many times by ÖKTAY and also earlier with other forms by other authors (KOSSWIG22, BREIDER23, RUST24, SCHWIER25, AIDA26).
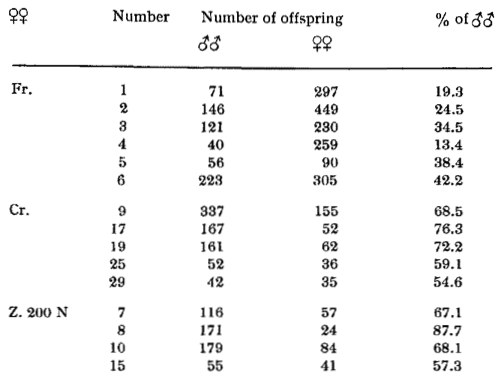 |
| Table I. X. helleri, progenies of different females, all paired with the same male (after BREIDER27) |
 |
| Table II. X. helleri, two females paired with different males, distinguishable from one another by the presence or absence of a dominant colour gene. The females were heterozygous for the dominant eolour gene Mo (after BREIDER27) |
- 6 M. GORDON, J. Hered. 37, 307 (1946).
- 14 W. BEERMANN, Biol. Zbl. 74, 525 (1955).
- 15 Ö. WINGE, C.R. Labor. Carlsberg Ser. phys. 21, 1 (1934).
- 16 Ö. WINGE, and E. DITLEVSEN, C.R. Labor. Carlsberg Ser. phys. 24, 227 (1948).
- 17 M. GORDON, Zoologica 36, 127 (1951).
- 18 Here it may be added that in the mammals principally the same type of localization of sex-deciding genes is present as in Chironomus or in the Cyprinodontid fishes. In the male sex all mammals bear a Y-chromosome. In the Y-chromosome — according to our present knowledge — the male determining gene (or genes) is localized. On the other hand the XO-type, which in Drosophila results in the production of males, yields females which at least in the mouse are completely normal and fertile. In man the same situation is principally true too.
- 19 M. ÖKTAY, Istanb. Üniv. Fen Fak. Mecm. B 24, 75 (1959).
- 20 M. ÖKTAY, Istanb. Üniv. Fen Fak. Mecm, B 24, 225 (1959).
- 21 H. BREIDER, Zool. Anz. 106, 277 (1934).
- 22 C. KOSSWIG, Z. ind. Abst.-Vererb.-lehre 57, 226 (1931).
- 23 H. BREIDER, Z. ind. Abst.-Vererb.-lehre 68, 265 (1935).
- 24 W. RUST, Z. ind. Abst,-Vererb.-lehre 79, 336 (1941).
- 25 H. SCHWIER, Z. ind. Abst.-Vererb.-lehre 77, 291 (1939).
- 26 T. AIDA, Genetics 21, 136 (1936}.
- 27 H. BREIDER, Z. wiss. Zool. (A) 146, 383 (1935).